Olá pessoa, tudo bem com vocês??
Hoje iremos abordar sobre a imunidade celular. Como vocês já viram falamos primeiramente sobre a inata, depois sobre os protagonistas (anticorpos, MHC, citocinas… TCR, BCR) da imunidade adaptativa, imunidade humoral e agora falaremos sobre a imunidade celular.
A imunidade celular como o próprio nome já diz é mediado por células, temos as principais TCD4 + e TCD8+ que irão atuar frente a patógenos diferentes, sendo direcionada pelos mesmos durante o processo de ativação.
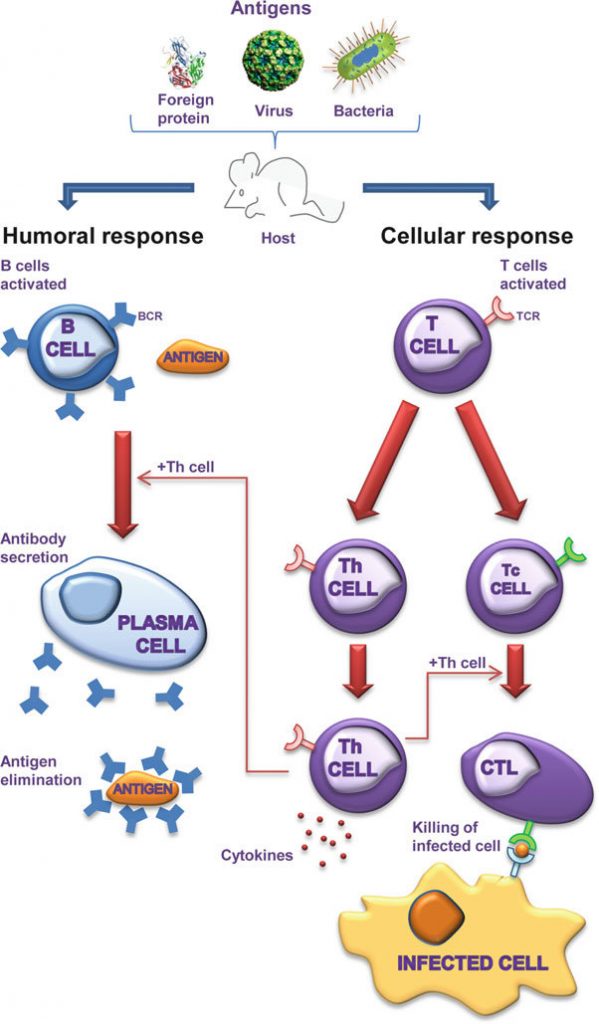
Comparação entre a resposta humoral e celular!
Então, vamos iniciar gerando um enredo para facilitar melhor esse entendimento pra vocês.
Nós temos um processo inflamatório acontecendo, todos aqueles mecanismos levaram a ativação das células dendríticas, as únicas capazes em interagir com as células T virgens e ativa-las.
Assim que as células dendríticas são ativadas uma cascata leva a o aparecimento de moléculas de MHC em sua superfície.
Caso o antígeno que a ativou tenha sido fagocitado as moléculas de MHC classe II que irão se ligar aos peptídeos formados que são os epítopos dessas proteínas (vale relembrar que tanto as moléculas de MHC quanto o TCR das células T só são capazes de reconhecer antígenos proteicos) que sofreram lise durante do processo de fagocitose.
Vamos entender o que acontece após?
Células dendríticas ativadas por antígenos extracelulares, fagocitados – MHC classe II
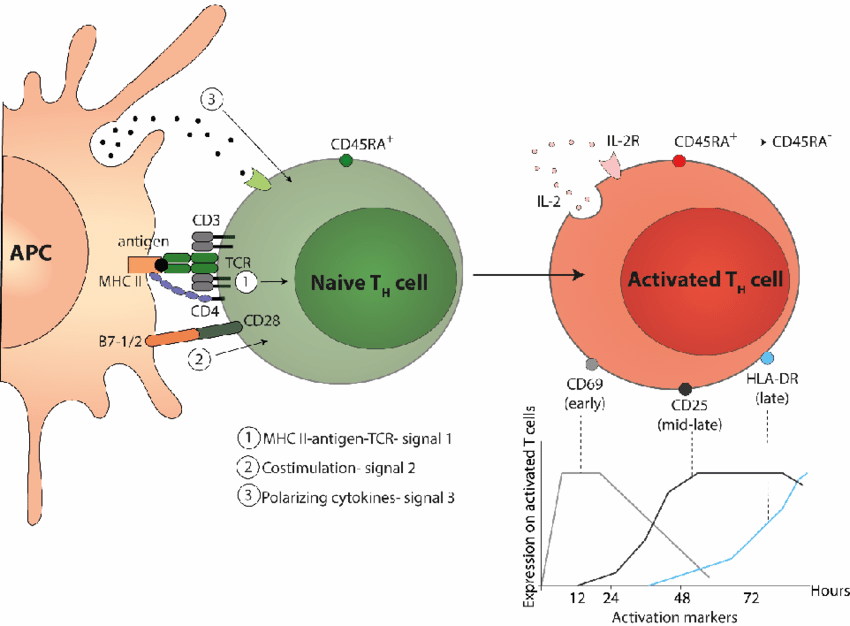
Após a ativação das células dendríticas ocorre a migração das mesmas em direção aos órgãos periféricos em busca de um linfócito T virgem. Ao chegar ao órgão periférico, à célula dendrítica faz o processamento do antígeno proteico e apresenta em sua superfície através de uma molécula de MHC.
Para que o linfócito T virgem possa ser ativado ocorrerá o reconhecimento dos peptídeos presentes na fenda da molécula de MHC através do receptor TCR, onde a região variável entrará em contato com o peptídeo preso no MHC, a cada três resíduos do peptídeo o TCR reconhecerá apenas um, sendo altamente específico e tendo capacidade de diferenciação por apenas um resíduo de aminoácido entre os microrganismos. Todo o complexo TCR formado pelas três proteínas CD3 e a Zeta ζ precisam ser estimulados para que de fato ocorra a ativação das células T.
Como dito acima, essa célula dendrítica foi ativada por um antígeno extracelular e em sua superfície se encontra o MHC de classe 2 que possui um correceptor CD4 que irá se ligar ao receptor correspondente nas células T virgem que irá direcionar a linhagem mais efetiva para eliminação do patógeno, que no caso será a célula TCD4+. Esse reconhecimento citado, funciona como o primeiro estímulo para ativação.
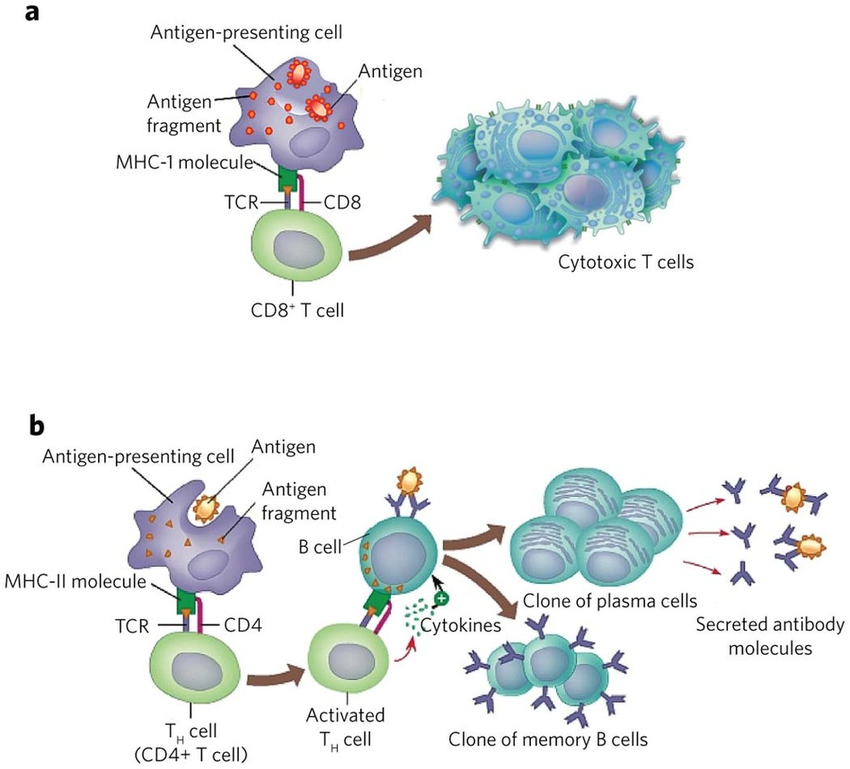
E caso o patógeno fosse intracelular, MHC classe I
A célula dendrítica iria processar no citosol esse antígeno que iria ativar as moléculas de MHC classe I que indo apara superfície com a presença do correceptor CD8 direcionaria a ativação do linfócito T virgem em linfócito TCD8+.
O que ocorre depois desse direcionamento?
Durante a ativação das dendríticas o patógeno promove paralelamente a ativação de coestimuladores (os quais serão o segundo estímulo) nessas células, aumento o número de receptores B7-1 e B7-2 os quais possuem receptores equivalente nos linfócitos T, os receptores CD28.
Com a interação desses receptores (B71 + o CD28) que funcionam como coestimuladores associados a ligação TCR + antígeno-MHC garantem a ativação de fato das células T. Sem a presença da coestimulação a ativação da célula T virgem em célula T efetora não acontecerá. Esses dois estímulos garantem que as células T sejam realmente ativadas frente à microrganismos e não a outras substâncias, o que explica a necessidade do uso de adjuvantes em formulações vacinais a base de proteínas purificadas ou patógenos inativados.
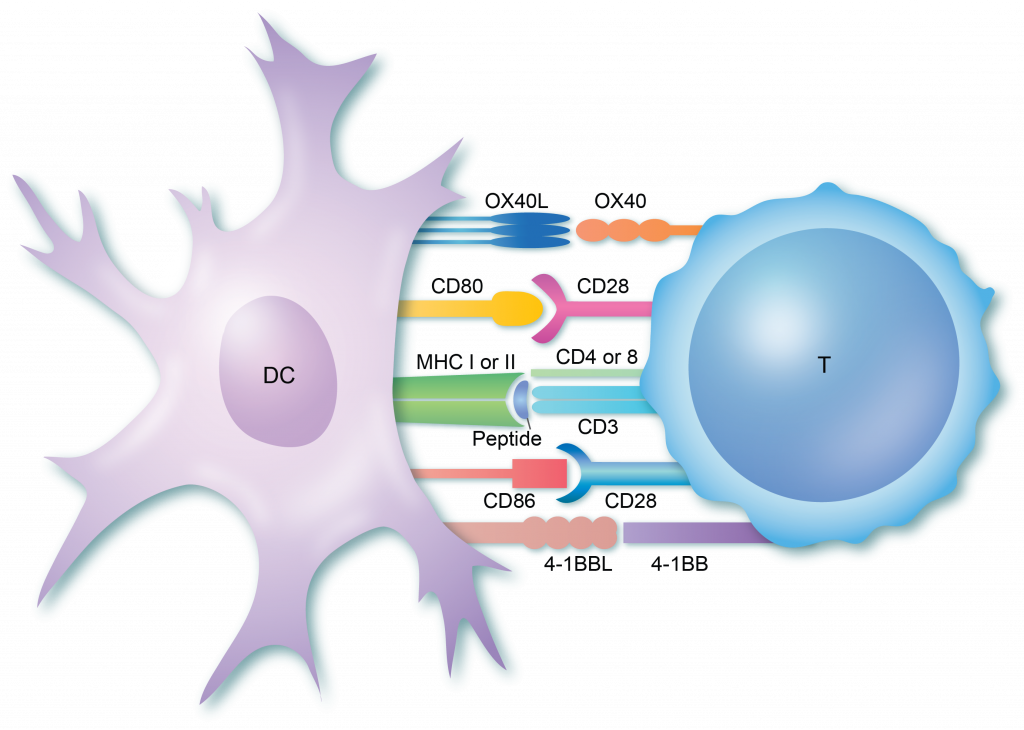
Outros fatores são importantes para que toda essa reação ocorra e de fato o linfócito T virgem seja ativado. A união entre o TCR e a molécula de MHC-antígeno possui baixa afinidade e com isso é necessário a presença de moléculas de adesão, como integrinas nos linfócitos T (antígeno 1 associado a função dos leucócitos – LFA-1) e um receptor intercelular 1 (ICAM -1) que irão promover a estabilidade da união dessas células para que toda a sinalização ocorra adequadamente e acarrete na ativação da resposta.
Assim, a ligação de outros receptores como CD40 entre essas células funcionará como uma retroalimentação para aumentar o estímulo B7/CD28 (promovendo a coestimulação e a cascata de eventos) levando a secreção de citocina IL-12 que intensifica o processo de diferenciação dessa célula.
Outros TCR serão estimulados, os receptores e demais proteínas de superfície formarão um aglomerado na superfície dessas células, formando a sinapse imunológica que é a região de contato entre a célula T e a APC, garantindo a ativação das células T.
Todos esses eventos tem por objetivo a transcrição de genes responsáveis pela expansão clonal dos linfócitos T específicos para antígenos e na diferenciação entre células efetoras e de memória, a primeira pode permanecer nos órgãos linfoides periféricos para eliminar células infectadas ou auxiliar os linfócitos B e outras podem migrar para o foco infeccioso para eliminar os patógenos.
As respostas proporcionadas pelos linfócitos T é através da produção de citocinas que podem atuar sobre elas mesmas (autócrinas) ou sobre outros células (parácrinas).
Assim que a célula T (principalmente a CD4+) é ativada ela secreta a IL-2 (dentro de 1 a 2 horas após ativação) que atua de maneira autócrina, as principais funções dessa citocina são de estimular a sobrevivência e a proliferação das células T, resultando no aumento do número das células T específicas do antígeno.
Os linfócitos T ativados começam a se proliferar clonalmente e no ápice do processo infeccioso (cerca de 1 semana) o número de células aumentam significativamente. A quantidade de células T CD8+ aumentam cerca de 10.000 vezes com o tempo estimado para dobrar sua população seja em cerca de 6 horas enquanto as células T CD4+ aumentam cerca de 100 a 1000 vezes.
Dentro de 1 ou 2 semanas após sua ativação, algumas células T expandidas foram diferenciadas em células efetoras e de memória e a maioria morrerá conforme os estímulos que iniciarão a resposta forem eliminados. O resultado é a contração dos clones expandidos e um retorno ao estado de equilíbrio inicial.
Diferenciação de células T virgens em células T efetoras
Parte das células T ativadas se diferenciam em células efetoras, as quais surgem cerca de 3 ou 4 dias pós contato com os microrganismos. Podendo ser CD4+ ou CD8+.
CD4+
Esse tipo de linhagem funciona ativando fagócitos e linfócitos B através da expressão de várias moléculas de superfície e secretando citocinas. Uma das principais proteínas expressas é a CD40L a qual se ligará ao receptor CD40 na superfície dessas células promovendo uma retroalimentação, que irá culminar na ativação de linfócitos B, macrófagos e células dendríticas.
Essas células podem se dividir em subgrupos de células efetoras, as quais produzirão tipos distintos de citocinas para cada tipo diferente de patógeno, apresentando uma resposta mais personalizada e específica.
Podem ser do tipo TH1, TH2 , TH9, TH17 e Reguladoras. A existência desses subgrupos mostra a atuação de algumas citocinas específicas por essas células, atuando para a eliminação de determinados patógenos. No entanto, deve-se saber que muitas células T CD4+ podem apresentam um mix de citocinas e, portanto, não podem ser classificadas em subgrupos, podendo existir plasticidade nestas populações, de modo que um subgrupo pode ser convertido em outro no decorrer da resposta imunológica.
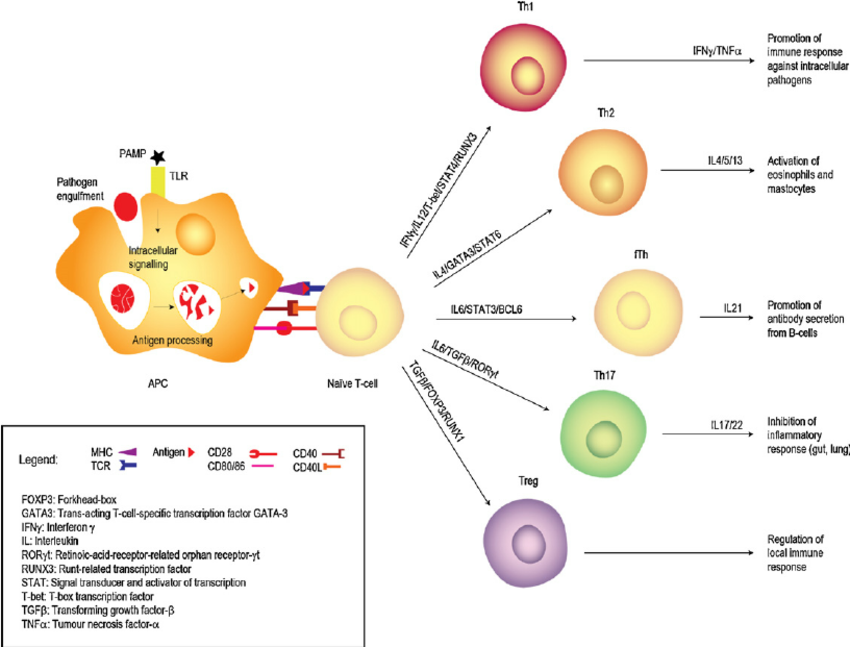
Subgrupo TH1
Estimulam o englobamento mediado por fagócitos e o extermínio de microrganismos, o componente-chave da imunidade mediada por células.
A citocina que é sintetizada principalmente nesse subgrupo é o interferon gama (IFN-ƴ). Potente ativador de macrófagos, especialmente na capacidade de matar micro-organismos ingeridos (denominado ativação clássica).
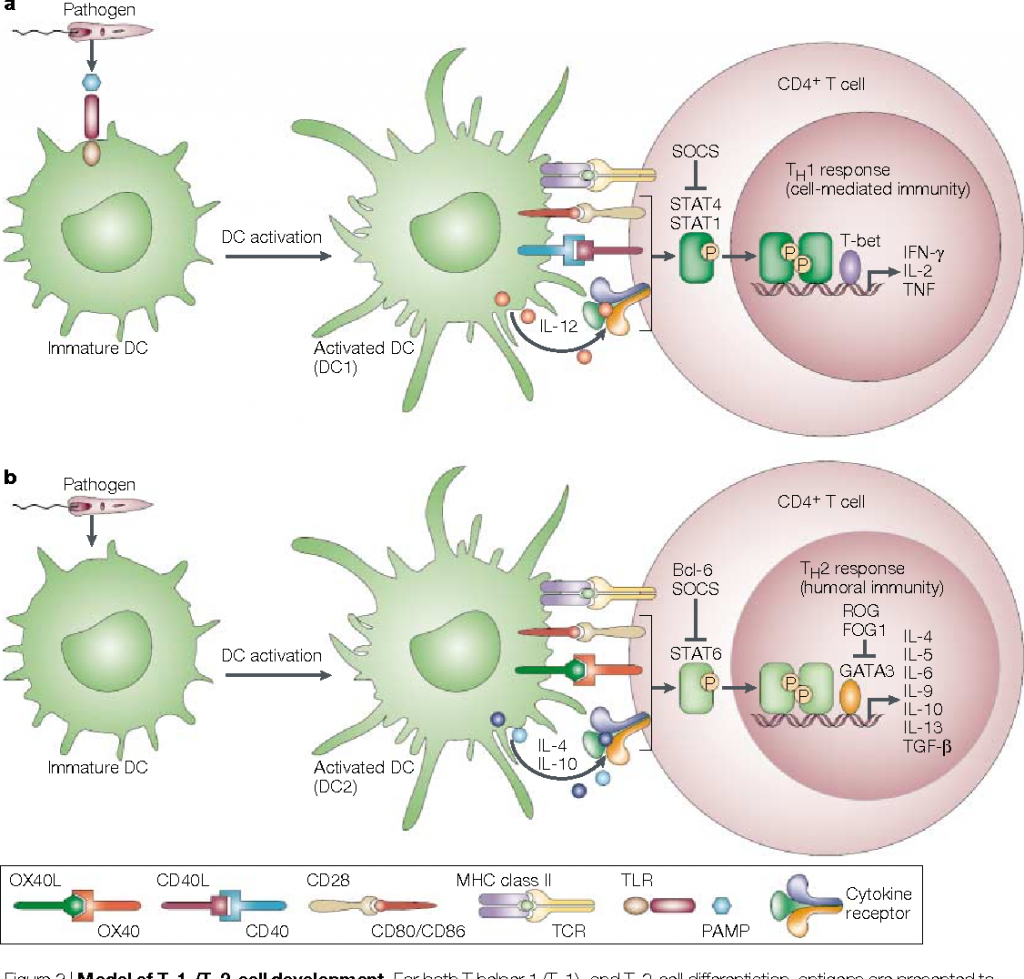
Também atuam na modificação da cadeia pesada dos anticorpos produzidos pelas plasmócitos, promovendo a fagocitose dos micro-organismos. Devido a essas ações esse tipo de resposta é importante para patógeno intracelular.
O IFN-ƴ também levam ao aumento da expressão das moléculas de MHC classe II e de co-estimuladores B7 na superfície de macrófagos e células dendríticas, amplificando a resposta por essas células.
Para que ocorra a diferenciação das células T CD4+ em TH1 é guiada pela presença de IL-12 e IFN-ƴ, em resposta a alguns patógenos as células dendríticas e os macrófagos produzem IL-12 enquanto as células NK produzem IFN-ƴ, deste modo quando as células T virgens são ativadas são expostas a estas duas citocinas, levando a diferenciação para células TH1, que produzirá IFN-ƴ, que não somente matará os micro-organismos intracelulares como também irá potencializar a diferenciação das células T em TH1 e vai inibir o desenvolvimento de TH2 e TH17.
Subgrupo TH2 e TH9
Estimulam a resposta independente de fagócitos mediada por eosinófilos, a qual é eficaz para parasitas (especialmente helmintos).
A citocina sintetizada nesse tipo de resposta é a IL-4 que estimula a produção de anticorpos IgE e também de IL-5 que estimula a atuação dos eosinófilos. A IgE ativa os mastócitos através da região Fc e se liga aos eosinófilos, essa interação é importante para a morte dos parasitas helmínticos.
Outra citocina expressa é a IL-13 que associada com a IL-4 promovem a expulsão dos parasitas dos órgãos mucosos e inibem a entrada de outros micro-organismos por meio da produção de muco.
A síntese dessas citocinas também ativa os macrófagos que são responsáveis pela reparação tecidual (denominada ativação alternativa). A atuação dessas citocinas também inibe a formação da resposta mediada pela TH1. Com isso, a eficácia da resposta imune mediada por células é o equilíbrio entre a resposta TH1 e TH2.
Para que ocorra a diferenciação para linhagem TH2 é necessário a presença de IL-4, no entanto sua produção é exclusiva pelas células T, como isso ocorre? Acredita-se que helmintos não promovem a atuação da IL-12 pela APC no momento do reconhecimento antigênico com isso a célula T sintetiza a IL-4 levando a diferenciação para a linhagem TH2. A outra fonte de IL-4 pode ser através dos mastócitos e eosinófilos ativados por esses patógenos.
Subgrupo TH17
As células TH17 induzem a inflamação, que funciona para destruir as bactérias extracelulares e fungos, e pode contribuir para várias doenças inflamatórias. Essas células secretam citocinas que recrutam leucócitos, principalmente os neutrófilos, para os locais de reconhecimento antigênico.
Essa subclasse foi descoberta no estudo em modelo animal para doenças de caráter inflamatório crônico como: esclerose múltipla, doença intestinal inflamatória e artrite reumatoide. Para que ocorra a diferenciação da célula T virgem em TH17 requer a atuação da IL-6 e IL-1 pelos macrófagos e células dendríticas e estas são expressas em resposta ao ataque de fungos e algumas bactérias.
CD8+
Esse tipo de células, conhecida também como citotóxica (CTL) expressam proteínas que entram pela membrana das células contaminadas e facilitam a entrada de enzimas que culminam no processo de apoptose dessa célula.
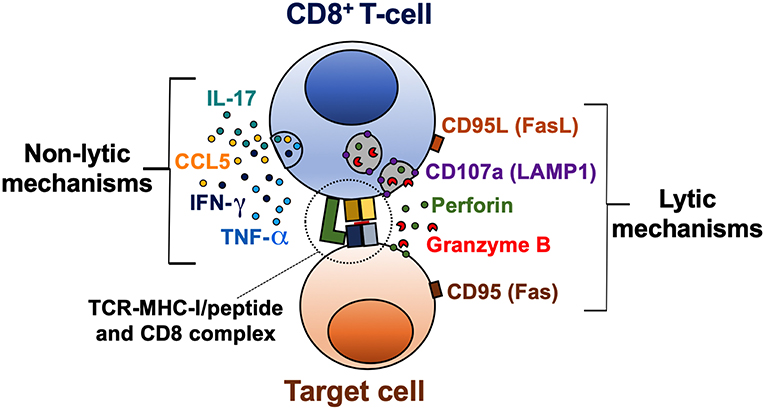
Linfócitos T de memória
Parte dos linfócitos T ativados seja CD4+ ou CD8+ serão diferenciados em linfócitos T de memória, estes sobrevivem após a infecção ser erradicada e o antígeno não estar mais presente. Elas ser de dois tipos, células de memória central e permanecem nos órgãos linfoides, ou podem ser células de memória efetoras e se encontram em tecidos periféricos, especialmente em barreiras mucosas e pele, e na circulação. Essas células necessitam da presença da citocina IL-17 para permanecerem vivas, não sintetizam citocinas, no entanto, na presença do antígeno elas voltam a atuar de forma rápida para eliminação do mesmo.
Mecanismos efetores
Dois tipos de reações mediadas por células estão presentes, se for mediada por linfócito T auxiliares CD4+, estes secretam citocinas que ativam os fagócitos a englobarem os micro-organismos e eliminá-los, se for mediada por linfócitos T citotóxicos CD8+ estes matam qualquer célula infectada contendo proteínas microbianas no citosol ou no núcleo, eliminando o reservatório infeccioso. Para que ocorra essa atuação, as células T virgens são ativadas nos órgãos linfoides como: linfonodos e baço, sofrem expansão clonal das células T antígeno-específicas, diferenciam-se na presenta das determinadas citocinas e podem ser efetoras ou de memória como já citado. Sendo efetora ela pode permanecer no órgão para eliminar possíveis células infectadas, para auxiliar os linfócitos B ou migram para o foco infeccioso. Em todos esses momentos ela precisará reconhecer os antígenos para poder atuar.

Conclusão
No artigo de hoje aprendemos sobre a resposta celular. A resposta celular é mediada pelos linfócitos T os quais, quando virgens, só podem ser ativados pelas células dendríticas previamente ativadas com a presença de moléculas de MHC em sua superfície. Quando a molécula de MHC é de classe I leva a formação de células TCD8+, quando a molécula de MHC é classe II levará a formação de células TCD4+. A ativação de fato dessas células só será feita através da união entre o MHC + TCR, coestimuladores e correceptores que permitam a sinapse imunológica por tempo suficiente para que a cascata de estímulos leve a transformações transcricionais.
Após essas ativações teremos células efetoras que atuarão de acordo com o direcionamento dessa resposta e teremos as células de memória que irão permitir que em um próximo contato esse antígeno/ patógeno seja eliminado mais rapidamente.
O sistema imune foi visto durantes essas oito semanas no intuito de trazer as bases para o conhecimento acerca dos mecanismos de ação do mesmo. Espero que vocês tenham curtido aprender ou reforçar um pouquinho dos seus conhecimentos sobre esse sistema que nos protege dia-a-dia!
Até a próxima!
Deixem seu comentário e compartilhem com os colegas!
Referências
ABBAS, A. K.; LICHTMAN, A. H. ; PILLAI, S. Imunologia celular e molecular. 6.ed. Rio de Janeiro: Elsevier, 2008.
ABBAS, AK; LICHTMAN, AH; PILLAI, S. Imunologia celular e molecular. 7° ed., Elsevier, 2012.
BENJAMINI, E.; COICO, R; SUNSHINE, G. Imunologia. 4. ed. Rio de Janeiro: Guanabara Koogan, 2002.
JANEWAY, C.; TRAVERS, P.; WALPORT, M.; SHLOMCHIK, M. J. Imunobiologia. O sistema imunológico na saúde e na doença. 7. ed. Porto Alegre: Artmed, 2010.
LIMA, F.A; Sampaio, M. C. O papel do timo no desenvolvimento do sistema imune. Pediatria, São Paulo, v. 29, n.1, p.33-42. 2007.
MARTÍNEZ, A.C.; ALVAREZ-MON, M. O sistema imunológico (I): Conceitos gerais, adaptação ao exercício físico e implicações clínicas. Rev Bras Med Esporte. v.5, n.3, 1999.